For det utrente øyet kan det se ut som om planter vokser ganske impulsivt, og plukker ut blader tilfeldig for å skape ett stort, grønt virvar. Ta imidlertid en nærmere titt, så vil du oppdage at noen få nysgjerrige vanlige mønstre dukker opp over hele den naturlige verdenen, fra den balanserte symmetrien til bambusskudd til de fascinerende spiralene til sukkulenter.
Disse mønstrene er faktisk konsistente nok til at kald, hard matematikk kan forutsi organisk vekst ganske godt. En antagelse som har vært sentral for studiet av phyllotaxis, eller bladmønstre, er at blader beskytter deres personlige rom. Basert på ideen om at allerede eksisterende blader har en hemmende innflytelse på nye, gir av et signal for å forhindre andre fra å vokse i nærheten, har forskere laget modeller som med hell kan gjenskape mange av naturens vanlige design. Den stadig fascinerende Fibonacci-sekvensen dukker for eksempel opp i alt fra solsikkefrøarrangementer til nautiluskall til kongler. Den nåværende konsensus er at bevegelsene til veksthormonet auxin og proteinene som transporterer det gjennom en plante er ansvarlige for slike mønstre.
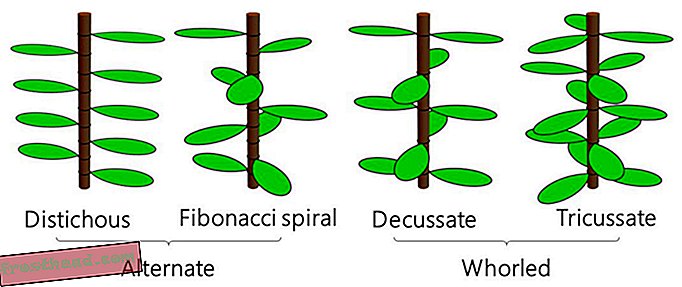 Bladarrangement med ett blad per node kalles alternativ phyllotaxis, mens arrangement med to eller flere blader per knute kalles whorled phyllotaxis. Vanlige alternative typer er distichous phyllotaxis (bambus) og Fibonacci spiral phyllotaxis (den saftige spiral aloe), og vanlige whorled typer er decussate phyllotaxis (basilikum eller mynte) og tricussate phyllotaxis ( Nerium oleander, noen ganger kjent som dogbane). (Takaaki Yonekura under CC-BY-ND)
Bladarrangement med ett blad per node kalles alternativ phyllotaxis, mens arrangement med to eller flere blader per knute kalles whorled phyllotaxis. Vanlige alternative typer er distichous phyllotaxis (bambus) og Fibonacci spiral phyllotaxis (den saftige spiral aloe), og vanlige whorled typer er decussate phyllotaxis (basilikum eller mynte) og tricussate phyllotaxis ( Nerium oleander, noen ganger kjent som dogbane). (Takaaki Yonekura under CC-BY-ND) Imidlertid fortsetter visse bladarrangementer å stamme populære modeller for plantevekst, inkludert Douady og Couder-ligningene (kjent som DC1 og DC2) som har dominert siden 1990-tallet. Et team ledet av University of Tokyo-forskere som studerer en busk kjent som Orixa japonica, fant ut at tidligere ligninger ikke kunne gjenskape plantens uvanlige struktur, så de bestemte seg for å revurdere modellen selv. Deres oppdaterte modell, beskrevet i en ny studie i PLOS Computational Biology, gjengir ikke bare det en gang unnvikende mønsteret, men det kan også beskrive andre, mer vanlige ordninger bedre enn tidligere ligninger, sier forfatterne.
"I de fleste planter har phyllotactic mønstre symmetri - spiral symmetri eller radial symmetri, " sier plantefysiologen University of Tokyo, Munetaka Sugiyama, seniorforfatter av den nye studien. ”Men i dette spesielle anlegget, Orixa japonica, er ikke det phyllotaktiske mønsteret symmetrisk, noe som er veldig interessant. For mer enn ti år siden kom en ide til meg om at noen endringer i den hemmende kraften til hvert blad primordium kan forklare dette særegne mønsteret. ”
Botanikere bruker divergensvinklene, eller vinklene mellom påfølgende blader, for å definere en plantes phyllotaxis. Mens de fleste bladoppstillingsmønstre holder en konstant divergensvinkel, vokser O. japonica- busken, som er hjemmehørende i Japan og andre deler av Øst-Asia, blader i en vekslende serie med fire gjentatte vinkler: 180 grader, 90 grader, 180 grader igjen, deretter 270 grader.
 En Orixa japonica busk med de forskjellige divergensvinklene på bladene synlige. (Qwert1234 via Wikicommons under CC BY-SA 4.0)
En Orixa japonica busk med de forskjellige divergensvinklene på bladene synlige. (Qwert1234 via Wikicommons under CC BY-SA 4.0) Dette mønsteret, som forskerne kalte “orixate” phyllotaxis, er ikke bare en engangs anomali, ettersom planter fra andre taxaer (som “rødglødende poker” -blomsten Kniphofia uvaria eller crepe myrtle Lagerstroemia indica ) veksler bladene i det samme komplisert sekvens. Fordi bladarrangementet dukker opp på forskjellige steder på det evolusjonære treet, konkluderte forfatterne at likheten kom fra en vanlig mekanisme som berettiget videre undersøkelse.
Etter å ha testet Douady- og Couder-ligningene med forskjellige parametere, kunne forfatterne produsere mønstre som var i nærheten av det vekslende orixatarrangementet, men ingen av de simulerte plantene stemte perfekt med O. japonica- prøvene de dissekerte og studerte. Så teamet bygde en ny modell ved å legge til en annen variabel i Douady og Couder-ligningene: bladalder. Tidligere modeller antok at bladenes hemmende kraft forble den samme over tid, men denne konstanten var "ikke naturlig fra biologiens synspunkt, " sier Sugiyama. I stedet tillot teamet av Sugiyama muligheten for at styrken til disse “holde seg unna” -signalene endret seg over tid.
De resulterende modellene - som teamet omtaler som utvidede Douady- og Couder-modeller, EDC1 og EDC2 - lyktes med å gjenskape, gjennom datastyrt vekst, de intrikate bladarrangementene til O. japonica . Utover denne bragden produserte de utvidede likningene også alle de andre vanlige løvmønstrene og spådde de naturlige frekvensene til disse variantene mer nøyaktig enn tidligere modeller. Spesielt når det gjelder spiralformede planter, spådde den nye EDC2-modellen "superdominansen" av Fibonacci-spiralen sammenlignet med andre arrangementer, mens tidligere modeller ikke klarte å forklare hvorfor denne spesielle formen ser ut til å vises overalt i naturen.
“Vår modell, EDC2, kan generere orixate mønstre i tillegg til alle hovedtyper av phyllotaxis. Dette er helt klart en fordel i forhold til den forrige modellen, sier Sugiyama. "EDC2 passer også bedre til den naturlige forekomsten av forskjellige mønstre."
 Blader på en Orixa japonica gren (øverst til venstre) og et skjematisk diagram av orixat phyllotaxis (til høyre). Det orixate mønsteret viser en særegen firesyklusendring av vinkelen mellom bladene. Et skannende elektronmikroskopbilde (midt og nede til venstre) viser vinterknoppen til O. japonica, der bladene først begynner å vokse. Primordialblader er merket sekvensielt med det eldste bladet som P8 og det yngste bladet som P1. Etiketten O markerer skuddens topp. (Takaaki Yonekura / Akitoshi Iwamoto / Munetaka Sugiyama under CC-BY)
Blader på en Orixa japonica gren (øverst til venstre) og et skjematisk diagram av orixat phyllotaxis (til høyre). Det orixate mønsteret viser en særegen firesyklusendring av vinkelen mellom bladene. Et skannende elektronmikroskopbilde (midt og nede til venstre) viser vinterknoppen til O. japonica, der bladene først begynner å vokse. Primordialblader er merket sekvensielt med det eldste bladet som P8 og det yngste bladet som P1. Etiketten O markerer skuddens topp. (Takaaki Yonekura / Akitoshi Iwamoto / Munetaka Sugiyama under CC-BY) Forfatterne kan foreløpig ikke konkludere med hva som nøyaktig får bladalderen til å påvirke disse vekstmønstrene, selv om Sugiyama spekulerer i at det kan ha å gjøre med endringer i auxintransportsystemet i løpet av en plantes utvikling.
Slike mysterier kunne løses ved "push and pull" mellom beregningsmodeller og laboratorieeksperimenter, sier Ciera Martinez, en beregningsbiolog som ikke var involvert i studien. Forfatterenes modell gir et spennende skritt mot en bedre forståelse av phyllotaxis og gir rom for andre botanikere å fylle ut hullene med planteseksjon og analyse.
"Med modeller, selv om vi kanskje ikke vet den nøyaktige mekanismen ennå, får vi i det minste kraftige ledetråder om hva vi skal se etter, " sier Martinez i en e-post. "Nå må vi bare se nærmere på molekylmekanismene i virkelige planter for å prøve å finne ut hva modellen spår."
 Et ovenfra-ned-utsnitt av bladoppstillingsmønstre i "orixate" phyllotaxis som nye blader (røde halvcirkler) dannes fra skudd-spissen (sentral svart sirkel) og vokser utover. (Takaaki Yonekura under CC-BY-ND)
Et ovenfra-ned-utsnitt av bladoppstillingsmønstre i "orixate" phyllotaxis som nye blader (røde halvcirkler) dannes fra skudd-spissen (sentral svart sirkel) og vokser utover. (Takaaki Yonekura under CC-BY-ND) Sugiyamas team jobber med å foredle modellen ytterligere og få den til å generere alle kjente phyllotactic mønstre. Et "mystisk" bladmønster, en spiral med en liten divergensvinkel, unngår fremdeles beregningsforutsigelse, selv om Sugiyama tror de er i nærheten av å knekke den løvrike koden.
"Vi tror ikke studien vår er praktisk nyttig for samfunnet, " sier Sugiyama. "Men vi håper at det vil bidra til vår forståelse av den symmetriske skjønnheten i naturen."